Como se vio en un Zapping anterior, primera parte de este artículo, el doctor Poole, de la Universidad de Estocolmo y otros colegas continúan buscando métodos para establecer la identidad, naturaleza y composición genética de
LUCA, antepasado común de todos los organismos terrestres, extintos o vivientes. LUCA figura en el árbol genealógico de todos nosotros, desde la más primitiva paleobacteria hasta Sir Isaac Newton.
LUCA no es un nombre, sino la sigla inglesa que significa "Último Ancestro Común Universal".
Dice Poole: "En un artículo aparecido hace un tiempo, Chuck Kurland ha criticado con firmeza la impaciencia de muchos científicos por atribuir a la evolución una transferencia horizontal masiva. Un aspecto particularmente interesante de su exposición es
que sugiere muchos factores anticientíficos que han contribuido a la moda de la transferencia horizontal. Su trabajo es en esencia un comentario acerca de cómo opera la mente científica, además de estudiar la transferencia genética".
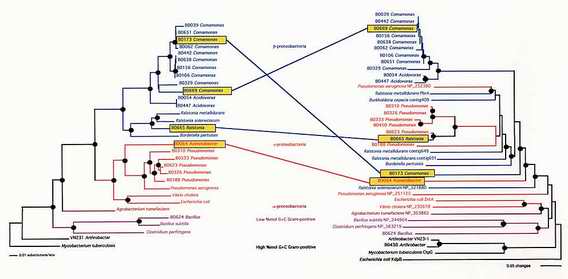
Ejemplos concretos de transferencia horizontal en bacterias
|
Las transferencias genéticas se encuentran justo en el límite de la sensibilidad de nuestras herramientas filogenéticas, y eso es lo que hace que las raíces del árbol evolutivo sean tan difíciles de encontrar y desenterrar. Tan es así, que el hecho de
que los investigadores no se hayan podido poner de acuerdo acerca de la forma del árbol no significa que eso sea consecuencia de una gran transferencia lateral.
En realidad, hay otra cuestión importante: la confiabilidad de los métodos que usamos para construir árboles evolutivos. Muchos sabios tienen una gran confianza en esos métodos, incluso siendo público y notorio que se basan en algoritmos matemáticos muy
cómodos pero que no necesariamente constituyen modelos reales de la secuencia de la evolución biológica.
Estos métodos parecen ser muy sólidos para los acontecimientos "recientes" de la evolución, y son ciertamente mucho más sólidos que los métodos que usamos para detectar transversalidad. El problema es que muy cerca de las raíces del árbol de la evolución,
los métodos pueden ser simplemente demasiado imprecisos y por lo tanto inútiles para investigar los eventos primigenios de la evolución. "En el peor de los casos, la situación sería como tratar de cronometrar a un atleta en los 100 metros llanos olímpicos...
¡utilizando un reloj de sol!", afirma Poole.
Según el investigador neocelandés residente en Suecia, los doctores David Penny y Bennet McComish han tratado de resolver este dilema investigando cuán atrás en el tiempo pueden proyectarse los modelos que se usan para reconstruir árboles evolutivos antes
de empezar a fallar. Sus conclusiones preliminares son que los modelos normalmente parecen trabajar un poco mejor de lo que les predice la teoría, pero que esos mismos modelos se equivocan de manera notable para eventos muy tempranos en la historia de
la evolución.
Mecanismos posibles de transferencia
|
|
También critican Penny y sus colaboradores la creciente tendencia a reportar árboles conflictivos como evidencia de la transferencia horizontal. Poole razona: "Dado lo difícil que parece ser reconstruir adecuadamente el árbol de la vida, es igualmente
difícil decir si dos respuestas contrarias son evidencia de transferencia, o si sólo están evidenciando las limitaciones de los métodos que usamos para reconstruir el árbol". Tiene razón: las pruebas de Penny sugieren que es irracional decir que hubo transferencia
transversal sólo porque dos árboles hechos a partir de dos genes distintos no señalan las mismas relaciones entre los organismos.
Escuchemos a Penny y McComish: "Si hablamos de divergencias muy antiguas, hay grandes problemas entre los conjuntos de datos de que disponemos. Es muy difícil entender por qué los investigadores muestran tanta confianza en los resultados, siendo que muchas
divergencias relativamente recientes entre mamíferos, aves o plantas con flores ni siquiera pueden ser resueltas hoy".
Los trabajos de Penny, y los de otros investigadores que nos han dado los resultados de árboles evolutivos a partir de genomas completos, parecen apoyar la afirmación de Kurland en el sentido de que "la transferencia horizontal es una ideología que pide
a gritos ser refutada".
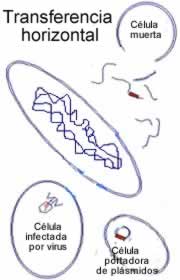
Sin embargo, la lateralidad existe, hasta cierto punto. De hecho, la tecnología biológico-industrial actual de transgenicidad es solamente la utilización por parte del Hombre de lo que ha sido dado en llamarse "ingeniería genética natural".
He aquí algunos de los mecanismos naturales de ingeniería genética que los laboratorios de biología molecular utilizan en forma rutinaria (tal como los describe Poole):
Plásmidos: pequeñas piezas de ADN, generalmente circulares, que a menudo contienen genes que los capacitan para trasladarse de una bacteria a otra.
Virus: muchos de ellos se introducen en el ADN de su víctima, y pueden ser modificados para llevar trozos de ADN "a medida".
Absorción —natural o asistida— de ADN ajeno por parte de células bacterianas.
Endonucleasas de restricción, enzimas que actúan como tijeras moleculares para cortar el ADN con toda precisión.
Sabemos también que es posible identificar transferencias de genes ocurridas en tiempos remotos, tan lejanas como dos mil millones de años atrás. Los biólogos pueden identificar de inmediato los genes de organismos eucariontes que han sido introducidas vía
mitocondrias, un órgano celular que es en realidad de origen bacteriano. Algunos de los genes de las bacterias que se introdujeron en las células y se convirtieron en mitocondrias residen ahora en el interior del núcleo.
Volviendo a la biología de los genes nómades, hay estudios que gozan de consenso y que dicen que debemos comenzar pensando que los genomas bacterianos (y tal vez los arqueanos también) fueron grupos de genes siempre cambiantes, pero solo hasta cierto grado.
Genes que son críticos para el funcionamiento de cualquier célula —llamados "genes ama de casa"— que forman la "sustancia" del genoma.
Genes que van y vienen haciendo al genoma más "flexible".
El genoma flexible podría ser una ventana al "pool genético nómada" de las bacterias. Lan y Reeves apuntaron en 2000 que las cepas de bacterias estrechamente relacionadas poseen genes que difieren hasta en un 20%, y esto obligará a la ciencia a reevaluar
cómo categorizamos especies diferentes. Se espera, sin embargo, que el genoma flexible pueda decirnos cómo vive en realidad una especie particular. La pérdida de genes no es particularmente sorprendente, en realidad, cuando se habla de especies que están
en una perpetua competencia para reproducirse tan rápido como pueden. La verdad es que los genes que no se utilizan no tienen por qué conservarse. La máxima de la selección natural es "úsalo o deshazte de él", y tenemos miles de ejemplos en la biología.
Nuestro apéndice y nuestro coxis van por ese camino, por ejemplo.
Este escenario de genoma de base y genoma flexible —si es correcto— es muy importante para la investigación de LUCA porque, en teoría, podremos recuperar muchas características "universales". Esto rige también para muchas transferencias horizontales primitivas
efectuadas a través de las mitocondrias. Podríamos ser optimistas en el sentido de que podremos analizar algunos patrones de transferencia horizontal de genes, aunque tengamos que ser cuidadosos al proyectarnos tan atrás en los tiempos remotos.
Cuando analizamos el proceso de transferencia horizontal, nos damos cuenta de que es una nimiedad si lo comparamos con lo que debió ocurrir en las etapas tempranas de la evolución de la célula eucarionte. La idea aceptada hoy en día es que la célula eucariota
se produjo tras una fusión masiva (llamada "transferencia final de genes") incluso entre bacterias y arqueas. Esto también presenta problemas para la biología de LUCA:
Los escenarios de la gran fusión desafían la división de los organismos vivos en tres reinos de Woese.
En vez de un árbol con tres ramas en cuya raíz está LUCA, la fusión tiene dos linajes de cuya unión surgen los eucariontes.
La fusión está en flagrante conflicto con el concepto de un enlace directo entre la biología eucariota y el mundo basado en el ARN.
Muchos investigadores han sostenido que los genes de un eucarionte promedio parecen ser una mezcla de genes de tipo bacteriano con genes de tipo arquea. Esto es lo mismo que decir que, a nivel genético, los eucariontes parecen ser una especie de mezcla
entre arqueas y bacterias.
Para comprender este fenómeno ha sido esencial dividir a los genes en dos categorías: "informativos" y "operativos". Los genes informativos son los que están involucrados en la copia, almacenamiento y regulación de la información genética, mientras que
los operativos contienen las recetas para producir proteínas para la síntesis y la catálisis de moléculas en el interior de la célula, y están enormemente presentes en el metabolismo de la energía.

Captura de ejemplares de arqueas en aguas termales
|
En consonancia con conocimientos anteriores, se ha demostrado recientemente que hubo necesidades y motivos para que se produjera la mezcla de genes arqueos y bacterianos en las células superiores, y que tal hecho puede rastrearse en la actualidad:
Los eucariotas y las arqueas comparten más genes informativos.
Los eucariotas y las bacterias comparten más genes operativos.
Alguien ha sugerido una buena analogía para lo que sucedió: una fusión corporativa. Dos compañías deben unirse en una sola (Eucariontes SA). En vez de duplicar todo y tener dos departamentos para cada aspecto de la nueva empresa, el gerente encargado de
la fusión conservó uno de cada uno, con el resultado de que el departamento Informaciones viene de Arquea y Cía., mientras que Operaciones proviene de Bacterias SRL.
El debate subsiguiente acerca de cuánta transferencia horizontal hay entre los organismos trajo consigo una de las contribuciones más importantes. Ella consiste en no estudiar los genes, sino las redes genéticas. Es asombrosa la manera en que Eörs Szathmáry
y sus colegas demostraron en 2001 que, aunque los genes operativos de los eucariontes parecen provenir de las bacterias, la estructura de las redes metabólicas producidas por esos mismos genes es parecidísima a la de las arqueas. Si seguimos con la metáfora
de las empresas, el gerente mantuvo el personal de Operaciones de Bacterias SRL pero adoptó los esquemas de trabajo de Arquea y Cía.
Es una idea emocionante, y no hay hoy en día discusión alguna acerca de que los eucariontes modernos son producto de la fusión de las dos ramas. Sin embargo, es difícil establecer qué significó exactamente aquella unión para el origen de la célula eucarionte.
Hay algunos temas en discusión:
¿Por qué ese tipo de fusión parece haber sucedido sólo una vez?
Nadie ha observado una fusión entre bacterias y arqueas modernas.
¿Por qué no existen "antieucariontes", es decir, organismos con los genes informativos de las bacterias y genes operativos de arqueas?
Muchas características presentes exclusivamente en los eucariontes son dificilísimas de explicar mediante una fusión.
En realidad, hay muchas maneras de explicar la fusión y, en consecuencia, existen opiniones diferentes acerca de cómo llegó a formarse la célula eucariótica.
Si los eucariotas son el resultado de la fusión entre una bacteria y una arquea, entonces la división en tres dominios apoyada en el trabajo de Carl Woese estaría equivocada. La fusión implicaría que cada cosa presente en la biología eucarionte es, o bien
una innovación reciente específica de ese dominio, o bien un remanente de la biología de arqueas y bacterias. En otras palabras, si usted quiere aprender sobre LUCA, los únicos lugares donde vale la pena buscar es en las arqueas y bacterias. A menudo se
asume esto —independientemente de la fusión— y es un tema contra el cual muchos científicos han protestado, Poole incluido.
Para comprender mejor las motivaciones de los variados escenarios de fusión que intentan explicar el origen de la célula eucariota y el impacto de los mismos en la biología "luquiana", conviene concentrarse en los hechos de fondo, en vez de perderse estudiando
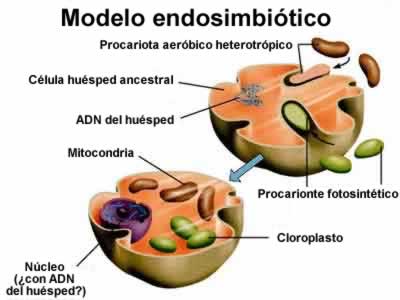
los detalles. En 1998, Laura Katz hizo un resumen de estos escenarios, aunque después de su trabajo aparecieron otros (entre 2000 y 2002). Las teorías de la fusión provienen de la "teoría endosimbiótica", que explica el origen de las mitocondrias.
La teoría endosimbiótica fue formulada por primera vez por Mereschkowsky a principios del siglo XX, pero fue actualizada y reintroducida por la bióloga Lynn Margulis en la década del ´70.
Esta teoría sostiene que la mitocondria, llamada a veces la "central de energía" de la célula, fue originalmente una bacteria que se "mudó" al interior de un ancestro de los eucariotas modernos.
Las similitudes tanto genéticas como estructurales han demostrado sin el más mínimo asomo de duda que la teoría endosimbiótica es correcta. El ADN de la mitocondria está estrechamente emparentado con el ADN bacterial,
y no tiene prácticamente nada en común con el ADN del núcleo de la célula eucarionte que lo alberga.
Se acepta hoy ampliamente el hecho de que este evento aconteció sólo una vez.
A pesar del gran consenso de que disfruta, hay actualmente un gran debate sobre la teoría de los endosimbiontes:
¿Cómo se formó exactamente esta "sociedad"? (Por ejemplo: ¿a partir de metabolismos basados en el oxígeno o en el hidrógeno?).
¿Qué era el organismo que originalmente "engulló" a la bacteria? ¿Un eucarionte? ¿Una arquea?
La primera pregunta abre una verdadera "Caja de Pandora" (que Poole evita en su trabajo), y es hoy una fuente de intenso debate. La segunda tiene un mayor impacto en la biología de LUCA, pero ambas han sido embrolladas innecesariamente. La realidad última
es que los datos sobre genes y redes genéticas que apoyan la teoría de la fusión entre arquea y bacteria, también sirven para apoyar una teoría de fusión entre eucariontes y bacterias.
Endosimbiosis, origen de las células superiores
|
La investigación se encuentra actualmente en este estado:
Todo el mundo coincide en que la mitocondria evolucionó a partir de un antepasado bacteriano (aunque el debate se centra hoy en cómo era ese antepasado bacteriano y en cómo interactuaba con su huésped).
Hay discusiones acerca de si ese huésped era eucarionte o arquea.
La fusión bacteria-arquea exige que todos los genes exclusivos de los eucariotas hayan evolucionado en tiempos postluquianos y postfusión. Es decir que no descienden directamente de LUCA.
Esto entra en conflicto con el concepto de un LUCA basado en ARN y con los tres reinos del árbol de la vida de Woese, ya que ambos necesitan que los eucariontes desciendan directamente de LUCA.
Entonces, ¿cómo distinguimos entre un huésped arqueo y uno eucariótico? La clave del asunto se divide en dos partes: una es histórica y la otra exige una consideración cuidadosa de los parentescos entre arqueas y eucariontes.
Los aspectos históricos se centran alrededor del cambio de escenario: del primero, con un eucariota primitivo haciendo de huésped hacia el segundo —ampliamente aceptado hoy— de un huésped arqueo recibiendo a la
mitocondria bacteriana. Este cambio de paisaje tiene mucho que ver con las ramas cambiantes del árbol evolutivo eucarionte.
La relación entre arqueas y eucariotas es definitoria para la idea que los investigadores se hacen de la evolución celular.
¿Por qué se volvió tan popular, entonces, la hipótesis de la fusión? En realidad, para oponerse a la interpretación "clásica" (enunciada por Tom Cavalier-Smith en 1987), que identificaba a un grupo disperso de eucariontes como los "eslabones perdidos",
los así llamados "arqueozoos", que parecen eucariontes pero no tienen mitocondrias. Esta hipótesis decía que los arqueozoos evolucionaron antes de la introducción de las mitocondrias en el linaje de las células superiores, y estuvo en boga durante muchos
años, hasta que fue descartada a favor de la fusión.
Hay crecientes evidencias de que todas las células eucariontes albergaron mitocondrias alguna vez.
Por ende, es probable que los arqueozoos sean en realidad eucariontes normales que perdieron sus mitocondrias, en vez de haber carecido siempre de ellas.
Por ejemplo, un grupo de arqueozoos llamados microsporidios ha sido recatalogado por los taxónomos: hoy se acepta que habían sido incorrectamente colocados muy abajo en el tronco del árbol eucarióntico. En vez de
ser eslabones perdidos en el origen de la célula eucarionte, han evolucionado en forma mucho más reciente.
Si los arqueozoos no son los eslabones perdidos, el origen de los eucariotas muy bien puede haber sido simultáneo con la endosimbiosis que originó las mitocondrias.
La conclusión de todo lo anterior es que todos los eucariontes tuvieron probablemente mitocondrias y que, sin los arqueozoos en la base del esquema, el único antepasado de todos ellos es la arquea. "Voilà!", exclama Poole: "Ya tenemos nuestra
fusión".
Plásmido con genes de dos especies
|
"El punto importante a tener en cuenta", explica el artículo de Poole, "acerca de la teoría de la fusión es que es una visión parcial, basada mayormente en los datos de la genética. Hay asimismo un gran número de diferencias de estructura entre eucariontes
y procariontes —arqueas y bacterias— que no pueden explicarse a través de la fusión". Sin embargo, la inconsistencia más grave es que el escenario provisto por los árboles no encaja con ni es el mismo que el producido por las relaciones entre arqueas
y eucariotas y entre bacterias y eucariotas:
La hipótesis de Margulis deriva del análisis de los árboles. Hay hoy un abrumador consenso acerca de que la rama mitocondrial se encuentra dentro del árbol bacteriano, específicamente en un subgrupo llamado protobacterias
alfa. También se descubrió un origen bacteriano para los cloroplastos (organoides donde tiene lugar la fotosíntesis de las plantas y otros eucariontes fotosintéticos)
Las comparaciones entre genes relevantes de eucariotas y arqueas también debieran llevar a esta conclusión, aunque las evidencias demuestren que arqueas y eucariontes pertenecen a reinos diferentes.
Hay grandes semejanzas entre esto y el tratamiento que se ha dado a los arqueozoos. Si no hay grupos modernos de arqueas que parezcan haberse separado del tronco del árbol de la vida antes de la aparición de los
eucariontes, ¿debemos insistir con la teoría de la fusión? ¡Se necesitó evidencia incontrastable para determinar el origen de la mitocondria!
Otro problema tiene que ver con los eslabones perdidos. Si la desaparición de los arqueozoos se utiliza para apoyar la fusión, es igualmente razonable argumentar por la tesis contraria con la misma base: no hay organismos
intermedios entre arqueas y eucariontes con mitocondrias. Por ejemplo, no vemos ejemplos de arqueas que contengan mitocondrias, ni de arqueas con estructuras de tipo nuclear en su interior.
Con más de 2.000 millones de años separando la divergencia de las arqueas y eucariontes, sería incorrecto pedir que el arquea de la fusión haya sido como las arqueas modernas. Hemos llegado a la raíz del problema: no hay ninguna obligación inherente de
que la evolución vaya dejando tras de sí series de intermediarios sólo para que nosotros podamos con comodidad reconstruir sus trayectorias, tanto en lo que respecta a las arqueas como a los eucariontes. Los puntos de llegada pueden llegar a ser muy diferentes
de los puntos de partida, y tenemos que observar un montón de puntos de llegada diferentes, ¡de los cuales sólo podemos especular acerca de donde comenzaron!
Mientras que la esencia de la hipótesis arqueozoica es probablemente un error, no debiese ser abandonada totalmente. La explicación del origen de la célula eucarióntica por fusión o por las ideas de Margulis adolece de formas intermedias, pero era de esperar.
Esta falta de organismos intermedios está de acuerdo con la frase de Stephen Jay Gould: "la evolución es como un monte lleno de arbustos, no como una escalera recta".
Muchos especialistas sostienen que los datos pueden ser reconciliados con la fusión, con la teoría endosimbiótica y con los tres reinos de Woese. En verdad, tanto Poole como David Penny han argumentado que la fusión es la teoría que peor explica
los datos experimentales. Por dar un ejemplo: la fusión no encaja con la hipótesis de que ciertas funciones eucariotas —que se han perdido en arqueas y bacterias— en realidad pueden rastrearse hasta LUCA.
Lo que realmente implica todo esto es que tanto arqueas como eucariontes deben haber tenido un antepasado común más reciente que el que comparten con las bacterias. Esto a menudo se afirma en los libros, pero no es del todo cierto, porque las relaciones
entre los tres grupos son todavía objeto de controversia.
Conclusiones
Estamos entrando en un emocionante período en el trabajo de descubrir la historia de LUCA: el campo ha sufrido una aceleración y un gran ensanchamiento causado por nuevas y mejores ideas aplicadas al problema.
Aceptar y reconocer los desafíos implícitos en la reconstrucción de árboles evolucionistas es un paso muy importante en la dirección correcta. Igual de importante es la idea de usar el período en que el mundo giraba
alrededor del ARN —en el origen de la vida terrestre— para establecer aspectos esenciales de la naturaleza de LUCA.
A pesar de la inundación de datos de genomas que se están publicando día a día, es difícil predecir si llegaremos algún día a recopilar una lista completa de genes que poseyó LUCA. Podemos aislar ciertas características,
pero cada una de ellas debe ser evaluada con un extremo cuidado. El estudio del genoma mínimo de Mushegian y Koonin lo demostró y, al fallar, nos da un fuerte llamado de atención. La mayoría de los investigadores han jugado con ideas como esta, y muchos
probablemente se sintieron defraudados (aunque luego tranquilizados) por el hecho de que Mushegian y Koonin se les hayan adelantado.
La transferencia horizontal de genes es probablemente un factor de confusión de esos esfuerzos, pero es preferible pecar por defecto en cuanto a la importancia de este fenómeno en la historia de la vida. El panorama
que surge de la investigación de genomas sugiere que no todos los genes se transfieren con la misma facilidad, y eso puede implicar que existe una ecología subyacente en la naturaleza de la transferencia genética.
La hipótesis de la fusión tiene importantes consecuencias para LUCA. Si es correcta, LUCA debe haber sido como una arquea y/o bacteria, porque las características exclusivas de los organismos eucariontes aún no habían
evolucionado.
Los tres dominios que surgen de los trabajos originales de Woese permiten que todas las características de los tres reinos sean rastreadas hacia atrás hasta LUCA. La hipótesis de la fusión, expresada en su forma
más estricta, no lo hace. Aunque se puede discutir la idea de que todas las arqueas modernas pueden haber ido perdiendo funciones o estructuras, sería diametralmente opuesta a la naturaleza de LUCA tal como la sugieren los fósiles de la biología de ARN.
En la actualidad, varios hechos que se daban por sentados comienzan a ser cuestionados de nuevo:
¿Hubo en el origen tres dominios o sólo dos, con un tercero que apareció por fusión?
¿Fue LUCA parecido a un procarionte, parecido a un eucarionte o una mezcla de ambos?
El código genético ¿es el único posible?
La evolución primitiva ¿se apoyaba en la transversalidad o en la herencia?
¿Hubo un solo LUCA, varios o muchos?
El doctor Anthony Poole afirma que "responder cada una de estas preguntas llenaría fácilmente un grueso libro, y cubrir cada aspecto de la biología de LUCA en un solo artículo es totalmente imposible. Para el observador casual, la luquiología parece un
campo enormemente desordenado, pero todos tienen su teoría predilecta. Esto puede resultar emocionante, frustrante, y muchas veces hacer equilibrio al borde del absurdo. Sin embargo, a pesar de todo es signo de un saludable debate". Muchos puntos de vista
y diferentes vías de aproximación al problema significan que estamos a un paso de descubrir importantísimas respuestas a algunas preguntas fundamentales sobre el origen de la vida...
Apéndice: Fabricación de proteínas
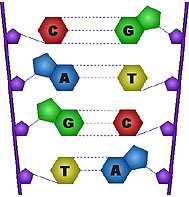
Correspondencia de las bases del ADN
|
|
Para hacer una proteína, la información almacenada en el lenguaje del ADN debe ser "traducida" al lenguaje de las proteínas en el cual, por ejemplo, en lugar de un alfabeto de 4 bases nitrogenadas se usa un alfabeto de 20 aminoácidos. La traducción es
primordialmente responsabilidad del ARN. Se hace una copia de trabajo del gen de ADN en soporte ARN, y esta copia se llama ARN mensajero (ARNm). El ARNm es leído por el ribosoma, que es la fábrica de proteínas que tienen todas las formas celulares de vida. La traducción
verdadera del lenguaje del ADN y el ARN al lenguaje proteico depende de otra forma de ARN llamada ARN de transferencia, ARNt.
El ARNt es la clave para entender la naturaleza real del código genético. En la construcción del ARN, cada una de las bases (A, C, G y U) se comporta como una palabra en sí misma.
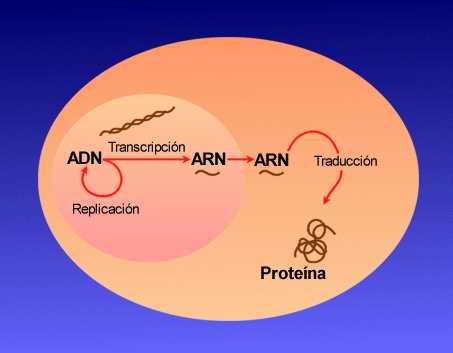
La síntesis proteica
|
Sin embargo, para traducir un alfabeto de 4 letras a uno de 20 (los aminoácidos
que componen la proteína), esas 4 palabras son agrupadas en palabras más largas, de tres bases cada una, llamadas "codones". Ello produce un "diccionario" de 64 palabras de tres letras cada una. De este modo, la célula representa cada aminoácido en este
código de "tripletes", con lugar incluso para agregar nuevas palabras. Se usan 61 de las 64 palabras, y las tres restantes significan "PARE DE CONSTRUIR LA PROTEÍNA", una señal de "fin del programa". Una importante característica de este código es su redundancia:
por ejemplo, hay seis codones diferentes que significan el aminoácido leucina. El sentido de esta redundancia es que el proceso de traducción sufre menos errores que si fuera un código unívoco ("uno a uno").
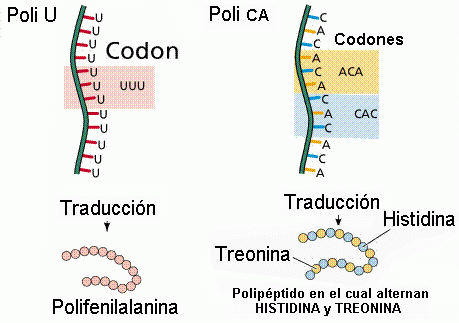
Dos ejemplos del proceso de traducción
|
La traducción requiere que cada ARNt se cargue con un aminoácido en un extremo. En el otro extremo, un "anticodón" encaja con el codón que codifica a ese aminoácido. La traducción necesita "amarrar" los ARNt cargados con su aminoácido al punto correcto
del ARNm a medida que el ribosoma se desplaza a través de la cadena, leyendo los tripletes uno por uno. Cuando el ARNt correcto (el que tiene el aminoácido que se necesita) está ubicado en su sitio, el ribosoma "pega" el aminoácido a la cadena proteica
en crecimiento. Todos los aminoácidos correctamente dispuestos forman por fin la proteína.
Más datos:
(Traducido, adaptado y ampliado por Marcelo Dos Santos de ActionBioScience y de otros sitios de Internet)